
















Apparto del Golgi
L’Apparato del Golgi fu scoperto alla fine dell’800 da Camillo Golgi in cellule nervose impregnate con acido osmico. In tale cellule risultava evidente un reticolo perinucleare che Camillo Golgi chiamò “apparato reticolare interno” (http://ppplab3.unipv.it/Pubblica/BioTec.pdf ). La presenza di tale apparato, variabilmente evidente in alti tipi cellulari, venne definitivamente confermata in tutte le cellule eucariotiche da immagini di microscopia elettronica. In onore del suo scopritore l’Apparto Reticolare Interno è stato denominato Apparato del Golgi.
Figura 1: Cellula nervosa impregnata con tetrossido di osmio. Le frecce indicano gli Apparati di Golgi, noti anche come pile golgiane e dittiosomi.
Figura 2: Cellule epiteliali del rene di opossum transgenici per una proteina che marca l’apparato di Golgi fusa con la GFP. Il segnale verde localizza il Golgi.
Da http://digitalimagingu.com/galleries/digitalvideo/spinningdisk/ok473laser/OK-EGFP-Golgi.html

Fig. 1

Fig. 2
Apparto del Golgi – morfologia
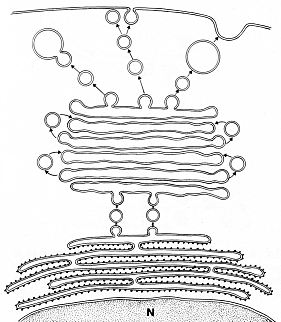
L’apparato del Golgi è costituito da una serie di cisterne (pile golgiane o, nelle cellule vegetali, dittiosiomi) ciascuna contenente un distintivo set di enzimi, in maggioranza glicosidasi. Le cisterne sono slargate in periferia, impilate l’una sull’altra e non intercomunicanti tra loro. Nelle pile golgiane si distinguono:
- faccia CIS o prossimale – contigua al Reticolo endoplasmatico, da cui riceve le microvescicole, le quali fondono tra a costituire una rete di vescicole interconnesse detta CIS Golgi Network (CGN).
- faccia trans o distale – lontana dal reticolo endoplasmatico – Dalla cisterna si staccano vescicole,interconnesse tra loro a costituire il Trans Golgi Network (TGN), da cui si staccano vescicole che diventano lisosomi, oppure vescicole di secrezione, o che si fondono con la membrana plasmatica
- cisterne mediane – Le cisterne interposte tra le CIS e TRANS.
Figura 1: Micrografia elettronica di una pila gogiana e sua schemtizzazione.
Immagine da http://porpax.bio.miami.edu/~cmallery/150/cells/c8.6×16.phagy.jpg
Figura 2: Schema dei diversi componenti di una pila golgiana

Fig. 1

Fig. 2
Apparto del Golgi – numero per cellula di pile golgiane
Il numero delle pile golgiane varia da uno (nei protozoi) a un centinaio (nelle alghe) e fino a mille ( nelle cellule gigante salivari degli insetti ditteri). In media sono presenti circa 20 pile golgiane per cellula; in ciascuna pila, in genere, sono presenti 4-5 cisterne.
Figura 1: Micrografia elettronica di una porzione citoplasmatica di una cellula algale. Osserva la presenza di numerosi dittiosomi (frecce).
Immagine da http://porpax.bio.miami.edu/~cmallery/150/cells/golgi2TEM.jpg
Figura 2: Ricostruzione 3D di una pila golgiana contenente 5 cisterne.

Fig. 1

Fig. 2
Apparto del Golgi – interconnessioni tra le pile golgiane
In ciascuna pila golgiana le cisterne non sono interconnesse. Tuttavia l’interconnessione può avvenire tra cisterne di pile golgiane contigue. Tali connessioni incrementano la velocità di transito delle proteine. La fusione tra le cisterne di pile differenti deve avvenire in modo rigorosamente corrispondente, ovvero che una cisterna di una pila, ad esempio la Cis, può fondersi solo con la corrispondente Cis dell’altra pila, per assicurare che sia mantenuto il controllo della direzionalità e della specificità dei processi che competono a ciascuna cisterna. Nel Golgi, quindi, anche in presenza di fusione tra differenti pile deve essere conservata l’ordinata progressione del processing enzimatico che compete a ciascuna cisterna.
In figura: Immagini che mostrano la continuità tra le cisterne di differenti pile golgiane in una cellula beta di un isolotto di Langherans di ratto. In alto le sezioni in Micrografia elettronica e sotto la ricostruzione tridimensionale.
Immagine da http://www.pnas.org/content/101/15/5565.full.pdf

Apparato del Golgi – traffico vescicolare – progressione vescicole
L’apparato del Golgi riceve, elabora e smista ad altri compartimenti il materiale (le glicoproteine) che riceve dal Reticolo endoplasmatico tramite microvescicole di trasporto. Nel Golgi l’elaborazione del materiale può avvenire secondo due modalità:
- progressione delle vesciole
- progressione delle cisterne
Secondo il modello della progressione delle vescicole il materiale, elaborato in un cisterna, viene in incluso in microvescicole di trasporto che gemmano lateralmente dai margini della cisterna per fondersi con la membrana della cisterna immediatamente successiva. Il processo continua fino alla cisterna TRANS. Dal TRANS si staccano vescicole che possono costituire i lisosomi primari (1) oppure le vescicole di secrezione (2) o anche fondersi con la membrana plasmatica (3). Nel modello della progressione delle vescicole si ha il movimento delle vescicole in direzione Cis-Trans. Tale movimento costituisce il “flusso vescicolare anterogrado“. Nel sito http://www.sumanasinc.com/webcontent/animations/content/vesiclebudding.html, è possibile osservare l’animazione dinamica del processo, nella versioni sia tappa per tappa sia continuata.
In figura modello modificato da: http://www.summagallicana.it/Volume2/008fig002.jpg

Apparato di Golgi: modello della progressione delle vescicole
Apparato del Golgi – traffico vescicolare- progressione delle cisterne
Il modello della progressione delle cisterne, valido soprattutto per le molecole di notevole dimensioni (quali ad esempio la cellulosa, la lattoalbumina), troppo grandi per essere contenute nelle micovescicole di trasporto, prevede che il materiale permanga nelle cisterne le quali avanzano maturando progressivamente in Cis, Mediana e Trans. Lateralmente, dalle cisterne gemmano delle microvescicole che trasportano nella cisterna precedente gli enzimi caratteristici di quella cisterna. Il materiale lascia il Golgi tramite vescicole si staccano dal TGN per formare i lisosomi, oppure le vescicole di secrezione o anche fondersi con la membrana plasmatica.
Secondo tale modello le cisterne, progredendo in direzione CIS-TRANS, realizzerebbero il Flusso delle cisterne anterogrado, mentre le vescicole, scorrendo nella direzione opposta TRANS-CIS , darebbero origine al flusso vescicolare retrogrado.
In figura: Modello della progressione delle cisterne.
Modificato da http://www.icampus.ucl.ac.be/courses/SBIM2520/document/genemol/pictures/golgi-origin.html
Coesistenza nelle cellule di pile golgiane a modalità sia della progressione delle cisterne sia della progressione delle vescicole.

Apparato del Golgi – traffico vescicolare – modalità combinata
E’ probabile che nelle cellule coesistano le due modalità di smistamento e elaborazione del materiale da parte delll’apparato di Golgi; ovvero che in una pila golgiana il materiale venga processato secondo il modello della progressione delle vescicole, mentre in una altra pila il materiale è elaborato secondo la modalità della progressione delle cisterne. A seconda del tipo cellulare e del materiale da elaborare una modalità può prevalere sull’altra.
In Figura: Apparato di Golgi – Modello della progressione delle vescicole.
Modificato da http://www.summagallicana.it/Volume2/008fig002.jpg

Apparato del Golgi – Riciclo vescicole RE-CIS Golgi
La gemmazione delle vescicole dal Reticolo endoplasmatico è mediato da specifiche proteine di rivestimento le COP-II, COat Proteins type II. Una volta che la vescicola si stacca dal RE le COP-II ritornano al RE.
Le vescicole arrivano al CIS- Golgi e scaricano il materiale. Dal CIS vengono riciclate al RE le vescicole che presentano sulla membrana una proteina con il segnale di ritenzione al RE. Tale segnale è costituito da 4 a.a. lisina, asparagina, glucina, leucina (K,D,E, L), il cui recettore è presente sulla membrana del RE. La gemmazione di tali vescicole è mediata dalle proteine di rivestimento del tipo COP-I.
Figura 1: Riciclaggio vescicole RE – CIS Golgi.
http://anatomy.iupui.edu/courses/histo_D502/D502f04/lecturef04/cell.f04/cellimages/Ross226.jpg
Figura 2: Riciclaggio delle vesicole contenenti il segnale di ritenzione al RE (KDEL)
http://www.steve.gb.com/images/science/er_golgi_trafficking.png

Fig. 1

Fig. 2
Apparato del Golgi – Riciclo vescicole RE-CIS Golgi
Le proteine COP-II rivestono solo le vescicole che gemmano dal Reticolo Endoplasmatico. Le proteine COP-I , invece, rivestano tutte le proteine che gemmano dalle cisterne golgiane, sia quelle del flusso vescicolare anterogrado sia quelle del flusso vescicolare retrogado.
Immagine modificata da: http://www.icampus.ucl.ac.be/courses/SBIM2520/document/genemol/pictures/golgi-origin.html

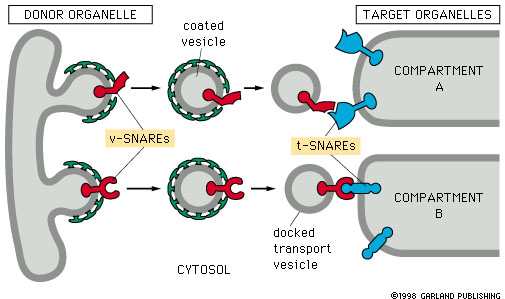
Apparato del Golgi – indirizzamento delle vescicole di trasporto
Le vescicole di trasporto presentano sulla membrana un particolare recettore per il materiale da trasportare. A ciascuno di tale recettore è associato dal lato citosolico una specifica proteina v-SNARE (vscicular-SNARE) che ne segnala il compartimento di destinazione. Su tale compartimento è presente una t-SNARE, che interagisce con la v-SNARE.
Coppie specifiche di proteine transmembarna v-SNARE e t-SNARE determinano la destinazione delle vescicole.
In figura: schema modificato da http://www.zoology.ubc.ca/~berger/B200sample/
unit_8_protein_processing/images_unit8/14_20.jpg

La destinazione delle vescicole di trasporto è mediata da coppie di proteine v-SNARE e t-SNARE
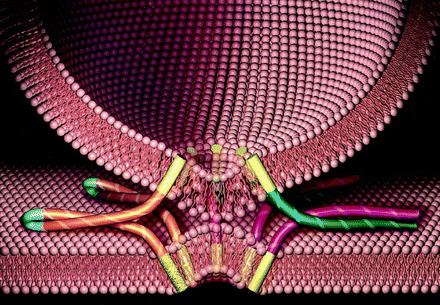
Apparato del Golgi – fusione delle vescicole di trasporto
La specifica segnalazione e fusione delle vescicole è medita dall’interazione di coppie complementari di proteine SNAREs: v-SNAREs (nella membrana della vescicola) e t-SNARE (nella membrana del comparto bersaglio). L’ adesione delle vescicole al comparto di destinazione è mediato dal legame tra le SNAREs, mentre la fusione delle vescicole e lo scarico del materiale nel comparto di destinazione è mediato da specifiche proteine di fusione (Rab e SNAPs,).
Figura 1: Il ruolo delle protiene SNARE nei processi dei seganlazione e fusione. lla segnalazione e The role of SNARE proteins in vesicle targeting and fusion.
http://mol-biol4masters.org/Co_and_Post_Translational_Events5-Cellular_Protein_Traffic_files/image073.jpg
Figura 2: Fusione tra le vescicole v-SNARE (in verde) e t- SNARE (in viola).
Da http://www.unil.ch/webdav/site/ib/shared/Mayer/Mayer.Fig1.jpg

Fig. 1

Fig. 2
Apparato del Golgi – maturazione delle proteine
Le cisterne golgiane differiscono sia strutturalemnte sia funzionalmente. Le membrane, infatti, presentano una differente composizione lipida e proteica che rendono ciascuna cisterna funzionalmente differenziata dalle altre. La componente proteica è costituita particolarmente da enzimi, glicosidasi e glicotrasferasi, che modificano l’ oligosaccaride, costituito da 10 residui zuccherini legati alle asparagine, che il Golgi riceve dal RE. Gli enzimi contenuti nelle cisterne conducono a due tipi due tipi di oligosaccaridi: oligosaccaridi complessi o oligosaccaridi ad alto contenuto di mannosio.

I due tipi principali di oligosaccaridi prodotti dal Golgi
Funzione dell’apparato del Golgi nella maturazione delle proteine
Le cisterne golgiane presentano uno specifico set di enzimi che modificano le glicoproteine durante il loro transito nelle cisterne in direzione Cis-Trans.
Nella immagine a fianco sono riportate le principali modifiche che in ciascuna cistena possono essere apportate ai residui oligosaccaridici.
In dipendenza delle modifiche apportate ai residui oligasaccaridi le glicoproteine vengono indirizzate ai lisosomi o alle vescicole di secrezione o alla membrana plasmatica.

Principali tappe della maturazione delle proteine nell'apparato di Golgi
Maturazione e smistamento delle proteine alla membrana plasmatica
Il modello proposto per le glicoproteine transmembrana della membrana plasmatica prevede che esse vengano sintetizzate nel RER. Il segnale di arresto blocca la traslocazione nel lume del RER, dove vengono glicosilate le asparagine della porzione sporgente nel lume. Dal RE si staccano le microvescicole con le glicoproteine transmembrana, che poi fondono con il Golgi. Nel transitare attraverso le cisterne gli oligosaccardi legati alle asparagine vanno incontro a modifiche, quali ad esempio:
- nel CIS l’eliminazione di residui di mannosio;
- nelle cisterne mediane l’aggiunta di N-acetilglucosammina;
- nel TRANS l’aggiunta di fucosio e acido sialico.
Dal TGN si staccano le vescicole con la componente glucidica delle glicoproteine transmembrana che sporgono nel lume delle vescicole. Dopo la fusione delle vescicole con la membrana plasmatica, le glicoproteine diventano proteine integrali della membrana plasmatica, con i residui zuccherini esposti solo nel versante extracellulare. Ciò spiega la esclusiva distribuzione della componente glucida delle glicoproteine o dei glicolipidi nel versante extracellulare della membrana plasmatica. Figura: http://porpax.bio.miami.edu/~cmallery/150/cells/sf5×8b.jpg

Schema della maturazione e smistamento delle glicoproteine alla membrana plasmatica
Maturazione e smistamento delle proteine alle vescicole di secrezione
Il modello proposto per la maturazione e smistamento delle proteine alle vescicole di secrezione prevede che esse siano sintetizzate e traslocate nel lume del RER, dove avviene la glicosilazione delle asparagine. Le glicoproteine poi passano all’apparato del Golgi. Nel transitare attraverso le cisterne gli oligosaccardi legati alle asparagine vanno incontro a modifiche:
- nel CIS l’ eliminazione di residui di mannosio;
- nelle cisterne mediane l’aggiunta di N-aceteliglucosammina;
- nel TRANS l’aggiunta di galattosio.
Dal TGN si staccano le vescicole che fondono con la membrana plasmatica e riversano nella matrice extracellulare le glicoproteine in esse contenute.
In figura: Modello della maturazione e smistamento delle proteine alle vescicolo di secrezione.
Modificato da http://porpax.bio.miami.edu/~cmallery/150/cells/sf5×8b.jpg

Secrezione costitutiva e secrezione regolata
Le proteine di secrezione vengono immesse in due tipi di vescicole di secrezione:
- Vesciscole non rivestite da clatrina ma da coatomeri che ininterrottamente gemmano dal TGN, fondono con la membrana plasmatica e riversano (Esocitosi) nella matrice extracellulare le glicoproteine contenute nel lume. Tale modalità di esocitosi rappresenta la “secrezione costitutiva” che è generalizzata in tutti i tipi cellulari, ad esempio i fibroblasti (esocitosi costitutiva di fibre collagene, la fibronectina, GAG, ect) plasmacellule (esocitosi costitutiva di immunoglobuline).
- Vescole rivestite da clatrina che si staccano dal TGN, si accumulano nel citoplasma e vengono successivamente rilasciate in seguito a specifici segnali provenienti dalla matrice extracellulare. Tale modalità di Esocitosi, denominata “secrezione regolata“, è tipica delle cellule ghiandolari endocrinie (secrezione regolata di ormoni) e soprattutto delle cellule degli epiteli ghiandolari esocrini ( esocitosi di secreti mucosi, sierosi, misti, ect) .
Da notare che la clatrina media anche la gemmazione delle vescicole contenenti le proteine lisosomiali.
In figura: Immagini modificate da ORGANITOS MEMBRANARES

Rappresentazione della secrezione costitutiva e regolata
Maturazione e smistamento delle proteine lisosomiali
Il modello proposto per la maturazione e smistamento delle proteine lisosomiali prevede che esse siano sintetizzate e traslocate nel lume del RER, dove avviene la glicosilazione delle asparagine. Le glicoproteine poi passano nella cisterna Cis dell’apparato del Golgi. In tale cisterna si ha la sforilazione dei residui del mannosio (M-6-P). Le proteine così marcate, senza subire altre modifiche, arrivano al TGN, dove sono riconosciute dal recettore del M-6-P ed incluse in vescicole rivestite da clatrina. Le vescicole successivamente trasportano le proteine lisosomiali ad un endosoma tardivo da cui gemmano le vescicole che riciclano al TGN i recettori del M-6-P.
In figura: Modello modificato da http://porpax.bio.miami.edu/~cmallery/150/cells/sf5×8b.jpg

Modello della maturazione e smistamento delle proteine lisosomiali
TGN centro di smistamento vescicolare
La gemmazione delle vescicole dal TGN è mediata da due tipi di proteine di rivestimento:
- Clatrina che media la gemmazione di vescicole della secrezione regolata o che contengono proteine lisosomiali.
- Coatomeri che mediano la gemmazione di vescicole della secrezione costitutiva.
Figura modificata da http://anatomy.iupui.edu/courses/histo_D502/D502f04/lecture.f04/cell.f04/cellf04.html

Schema delle vescicole che gemmano dal TGN
L’immagine qui riprodotta riassume i vari tipi di proteine di rivestimento implicate nel traffico vescicolare tra il RE, l’apparato del Golgi e la membrana plasmatica.
In figura: La via della secrezione – Ruolo delle proteine di rivestimento, COPI, CPOII e Clatrina.
Immagine da: www.uhmc.sunysb.edu/som/courses/mgac/Lect.%2018%20Transport.ppt

LISOSOMI
I lisosomi furono scoperti dapprima biochimicamente da De Duve su celllule di ratto, da cui riuscì ad isolare ed purificare i lisosomi attraverso delle semplici tecniche di centrifugazione. De Duve osservò che nella frazione lisosomiale l’attività delle idrolasi acide da lui studiati (fosfatasi acida, DNasi, RNAsi, glucosidasi, ect) era molto alta nel sedimento e bassa nel supernatante quando utilizzava metodi di isolamento dolci (preservanti le membrane cellulari). Tuttavia, dopo il congelamento della frazione lisosomiale o il suo isolamento con metodi drastici (destabilizzanti le membrane cellulari) l’attività delle idrolasi risultava alta nel supertante e bassa nel sedimento. Da tali esperimenti De Duve dedusse che le idrolasi acide dovevano essere contenute in organelli delimitati membrana che furono denominati “lisosomi” in base alle definizione data da Duve a tali organelli: “This Small Particle acts as the digestive tract of the living cell. Its enzymes dissolve the substances ingested by the cell and under certain circumstances can dissolve the cell itself”.

Esperimenti di De Duve sulla attività delle idrolasi acide
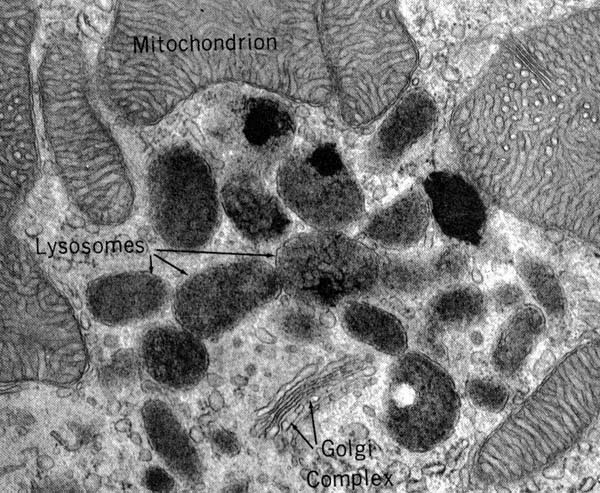
LISOSOMI – morfologia
Le indagini di microscopia elettronica confermarono le evidenze biochimiche di De Duve dimostrando che le idrolasi acide sono contenute in organelli citoplasmatici elettrondensi, delimitati da una membrana propria, dal diametro variabile da 50 nanometri a 5 mm. Successive indagini dimostrarono la presenza dei lisosomi in tutte le cellule eucariotiche.
Il set degli enzimi lisosomiali (proteasi, fosfatasi, lipasi, solfatasi, glicosidasi, RNasi e DNasi) può variare a secondo del tipo di cellula e dell’organismo considerato, in tutti i casi, però, è sempre presente la fosfatasi acida, che è quindi l’enzima marcatore dei lisosomi.
In figura: Micrografia elettronica con vescicole lisosomiali.
http://www.vialattea.net/spaw/image/biologia/lysosome.jpg

Genesi dei Lisosomi
Le proteine lisosomiali vengono sintetizzati dal RER, che ne opera la glicosilazione a livello delle delle asparagine. Nella cisterna Cis dell’apparato del Golgi i residui di mannosio delle proteine lisosomiali vengono fosforilati (M-6-P). Le proteine lisosomiali così marcate nel TGN vengono riconosciute dal recettore del M-6-P ed incluse in vescicole rivestite da clatrina. Le vescicole che si staccano dal TGN costituiscono i lisosomi primari. Tali organelli sono fortemente elettrondensi. Le idrolasi non digeriscono i lisosomi primari per vari motivi, tra cui:
- pH circa 6,5 (quello ottimale è pH 3- 5)
- la scarsità di acqua disponibile
- la membrana è resistente all’azione delle idrolasi grazie a specifiche glicoproteine transmembrana che rivestono la sua superficie interna.
- il fosforo legato al mannosio inattiva tutte le idrolasi, eccetto la fosfatasi che tuttavia non può agire per la scarsità di acqua e per il pH non ottimale.
Quando i lisosomi primari si fondono con gli endosomi tardivi diventano endolisosomi. Qui avviene il riciclaggio dei recettori del M-6-P mentre le idrolasi lisosomiali trovano le condizioni ottimali per la loro azione (l’acqua e il pH tra 3 e 5).
In figura: Modello Modificato da http://porpax.bio.miami.edu/~cmallery/150/cells/sf5×8b.jpg

Maturazione e smistamento delle proteine lisosomiali e strutture dei lisomi primari
Nell’immagine qui riportata vengono mostrati i dettagli della maturazione delle proteine lisosomiali, che comprendono: la marcatura nel Cis Golgi del mannosio 6 fosfato (M6P), il legame al recettore del M6P nel TGN, la gemmazione delle vescicole lisosomiali mediate dalla clatrina, la traslocazione delle proteine lisosomiali all’endosoma tardivo, dove avviene il riciclaggio dei recettori del M6P e l’attivazione delle proteine lisosomiali tramite la rimozione del M6P.
In figura: Maturazione delle proteine lisosomiali. Immagine modificata da www.uhmc.sunysb.edu/som/courses/mgac/Lect.%2018%20Transport.ppt



Le lezioni del Corso
1. Livelli di organizzazione in Biologia - Storia della citologia
2. Principi e tecniche in microscopia
3. Allestimento di preparati biologici in microscopia ottica ed elettronica
4. Composizione chimica della cellula
5. Principali metodi istochimicici, immuno-istochimici, colture in vitro, autoradiografia
6. Sguardo d'insieme ai virus, alla cellula procariotica ed eucariotica
8. Fisiologia della membrana plasmatica
9. Citoscheletro: generalità e i microfilamenti
10. Citoscheletro: microtubuli e filamenti intermedi
11. Giunzioni cellulari e adesione cellulare
12. Ribosomi, smistamento proteine, reticolo endoplasmatico
13. Apparato di Golgi, Lisosomi
I materiali di supporto della lezione
Consultare i capitoli dedicati all'apparato del Golgi e ai lisosomi nei testi di biologia cellulare e molecolare, quali ad esempio:
Karp. Biologia cellulare e molecolare (3a Edizione). Edises Editore
Becker et al. “Il mondo della Cellula”. Edises editore
Alberts et al. Biologia molecolare della cellula. Zanichelli Editore







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}